Verfasst vom Qubic Scientific Team
 Neuraxon Intelligence Academy — Band 3
Neuraxon Intelligence Academy — Band 3

1. Neuromodulation im Gehirn: Die Grundlage der adaptiven Intelligenz
Neuromodulation bezieht sich auf die Menge an Mechanismen, die regulieren, wie das Nervensystem zu einem bestimmten Zeitpunkt funktioniert, ohne seine grundlegende Architektur zu verändern. Dank der Neuromodulation kann das Gehirn schnell oder langsam lernen, explorativ oder konservativ werden und offen für Neuheiten bleiben oder sich auf das konzentrieren, was bereits bekannt ist. Die Verdrahtung ändert sich nicht; was sich ändert, ist die Art und Weise, wie diese Verdrahtung genutzt wird. Dieses Konzept ist zentral für das Verständnis von gehirninspirierten KI und der Architektur hinter Qubics Neuraxon.
Ionotrope vs. metabotrope Rezeptoren: Zwei Zeitskalen neuronaler Signalgebung
Um Neuromodulation richtig zu verstehen, ist es wichtig, zwischen zwei Formen chemischer Aktionen im Gehirn zu unterscheiden. Einerseits gibt es Neurotransmitter, die auf ionotrope Rezeptoren wirken, wie Glutamat und GABA. Diese Rezeptoren sind Ionenkanäle: Wenn sie aktiviert werden, erzeugen sie sofortige elektrische Veränderungen im Neuron, im Bereich von Millisekunden. Dies entspricht dem schnellen Niveau neuronaler Berechnung: konkrete Informationen werden übertragen, sensorische Signale integriert, schnelle Entscheidungen getroffen und die neuronale Aktivität erzeugt, die Wahrnehmung, Bewegung und Echtzeitgedanken stützt.
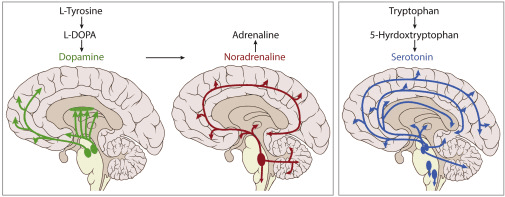
Andererseits gibt es Neurotransmitter wie Dopamin, Noradrenalin, Serotonin und Acetylcholin, deren primäre Wirkung über metabotrope Rezeptoren ausgeübt wird. Diese Rezeptoren erzeugen nicht direkt ein elektrisches Signal. Vielmehr aktivieren sie intrazelluläre Signalkaskaden, die die inneren Eigenschaften des Neurons über längere Zeiträume, Sekunden, Minuten oder mehr, verändern. Dies stellt das langsame dynamische Niveau der neuronalen Verarbeitung dar, das grundlegend dafür ist, wie das Gehirn sich anpasst und lernt.
Eine intuitive Art, über diesen Unterschied nachzudenken, ist durch die Metapher eines Seehafens. Ionotrope Rezeptoren sind wie Schwimmer, Surfer oder kleine Boote, die schnell ein- und ausfahren. Metabotrope Rezeptoren hingegen sind wie große Containerschiffe. Damit sie anlegen können, sind Genehmigungen erforderlich, Koordination notwendig und die Logistik des Hafens muss angepasst werden. Diese metabotropen Rezeptoren verändern die synaptische Plastizität und die Leichtigkeit, mit der ein Neuron reagiert – diese langsame Modulation überträgt keine Informationen, sondern verändert die internen Regeln des Systems.
Die vier Neuromodulatoren: Dopamin, Noradrenalin, Serotonin und Acetylcholin
Hier kommen die großen neuromodulatorischen Systeme ins Spiel. Jeder dieser vier Neurotransmitter spielt eine eigene Rolle bei der Regulierung, wie das Gehirn Informationen verarbeitet, lernt und sich anpasst:
Dopamin, das hauptsächlich aus dem ventralen tegmentalen Bereich und der substantia nigra stammt, signalisiert nicht unbedingt Freude, sondern wenn etwas für das Lernen relevant ist. Es passt die Sensitivität des Systems für Fehler und Neuheit an. Wie Schultz (2016) in seiner grundlegenden Arbeit zur Dopamin-Belohnungsprognosefehlerkodierung gezeigt hat, signalisiert Dopamin den Unterschied zwischen erwarteten und tatsächlichen Ergebnissen, ein Mechanismus, der für das Verstärkungslernen sowohl in biologischen als auch in künstlichen Systemen entscheidend ist.
Noradrenalin (Norepinephrin), das hauptsächlich aus dem Locus coeruleus freigesetzt wird, reguliert Erregung und das Gleichgewicht zwischen Erkundung und Ausbeutung. Wenn sein Ton hoch ist, wird das Gehirn empfindlicher für unerwartete Veränderungen und weniger an Routinen gebunden. Dies steht im Einklang mit der integrativen Theorie, die von Aston-Jones & Cohen (2005) vorgeschlagen wurde, die die Funktion von Locus coeruleus–Norepinephrin mit adaptiver Gewinnkontrolle und Entscheidungsfindung unter Unsicherheit verknüpft.
Serotonin, das in den Raphe-Kernen entsteht, moduliert Stimmung, Schlaf, Hemmung und Verhaltensstabilität. Wie in Dayan & Huys (2009) untersucht, drängt Serotonin das System nicht dazu, schnell zu lernen, sondern eher abzuwarten, impulsive Reaktionen zu vermeiden und das Verhalten aufrechtzuerhalten, wenn die Umgebung unsicher ist. Es spielt eine entscheidende Rolle für Geduld und langfristige Planung.
Acetylcholin, das aus den Basalganglien im Hirnstamm freigesetzt wird, spielt eine zentrale Rolle bei Aufmerksamkeit und kontextabhängigem Lernen. Es erleichtert das Öffnen kortikaler Netzwerke für relevante sensorische Informationen und ermöglicht synaptische Plastizität, wenn die Umgebung dies erfordert. Es ist besonders wichtig, wenn etwas Neues gelernt werden muss, was es entscheidend für adaptive neuronale Berechnung macht.
Dank dieser kombinierten Wirkung kann derselbe Reiz je nach neuromodulatorischem Zustand unterschiedliche Reaktionen hervorrufen. Der Schaltkreis ist derselbe, aber die Art und Weise, wie er funktioniert, hat sich geändert. Deshalb reagiert das Gehirn nicht auf die gleiche Weise, wenn es aufmerksam ist, wie wenn es müde ist, noch lernt es auf die gleiche Weise in routinemäßigen Situationen wie im Angesicht von Neuheit oder Überraschung.
Die Metaebene: Fenster der Plastizität und adaptives Lernen
Es gibt auch eine dritte, tiefere Ebene, die als Metaebene der neuronalen Regulation verstanden werden kann. Diese Ebene reguliert nicht direkt die neuronale Aktivität oder deren Geschwindigkeit, sondern die Bedingungen, unter denen das System sich nachhaltig ändern kann. Im Gehirn garantiert die gleichzeitige Aktivität zwischen Neuronen nicht unbedingt Lernen. Damit eine Verbindung gestärkt oder geschwächt werden kann, muss der neuromodulatorische Zustand dies erlauben. Es ist, als gäbe es ein stilles Signal, das sagt: „jetzt ja“ oder „jetzt nicht“.
Neuromodulation wirkt somit als ein System, das Fenster der Plastizität öffnet oder schließt und entscheidet, wann ein Fehler, eine Erfahrung oder eine Zufälligkeit konsolidiert werden sollte. Diese multiskalare Architektur, schnell, langsam und meta, existiert, weil ein intelligentes System nicht immer die gleichen Regeln anwenden kann. Wie Marder (2012) in ihrer wegweisenden Rezension erklärte, ist die Neuromodulation neuronaler Schaltkreise, wie das Gehirn Verhaltensflexibilität erreicht, ohne seine Architektur neu zu bauen.
Der Zustand des Körpers, Energielevels, Müdigkeit oder Schmerz sind Teil der internen Umgebung. Neuheit, Bedrohung, Gelegenheit, Wiederholung oder Vorhersehbarkeit sind Teil der externen Umgebung. Neuromodulatorische Systeme übersetzen diese Bedingungen in funktionale Zustände. Durch Dopamin, Noradrenalin, Serotonin und Acetylcholin bewertet das Gehirn, ob eine Situation Lernen verdient, ob Vorsicht erforderlich ist, ob Erkundung oder Erhaltung vorzuziehen ist und ob ein Fehler informativ oder lediglich Rauschen ist. Die Umgebung diktiert nicht direkt die Antwort, sondern moduliert die Regeln, nach denen das Gehirn reagiert. Dieses Prinzip steht im Mittelpunkt dessen, was Friston (2010) als das Prinzip der freien Energie beschrieb, ein einheitlicher Rahmen, der darauf hindeutet, dass das Gehirn kontinuierlich Überraschungen durch adaptive interne Modelle minimiert.
2. Warum große Sprachmodelle und transformerbasierte Architekturen an Neuromodulation fehlen
Große Sprachmodelle (LLMs) und transformerbasierte Architekturen besitzen keine Neuromodulation. Obwohl sie lange Sequenzen verarbeiten und bemerkenswerte Leistungen in der Verarbeitung natürlicher Sprache erzielt haben, fehlt ihnen ein System, das das Betriebsregime des Modells während der Inferenz dynamisch reguliert.
Die statische Natur von transformerbasierten KI-Systemen
Das Lernen in LLMs erfolgt während Trainingsphasen, die völlig getrennt von der Anwendung sind. Gewichte werden durch Rückpropagation des Fehlers angepasst, und sobald das Training abgeschlossen ist, befindet sich das Modell in einem festen Zustand. Während der Inferenz gibt es keine Plastizität und keine dauerhafte Veränderung als Funktion des Kontexts. Das System entscheidet nicht, wann es angemessen ist zu lernen und wann es stabilisieren sollte, weil es nicht lernt, während es arbeitet. Dies ist die grundlegende Einschränkung, die die jüngste Forschung bestätigt hat: LLMs fehlen echte interne Weltmodelle und die Fähigkeit, in Echtzeit anzupassen.
Einige von Neuromodulation inspirierten Ansätze versuchen, bestimmte Effekte zu approximieren, indem sie Parameter wie die Lernrate während des Trainings anpassen, Subnetzwerke aktivieren oder deaktivieren oder Aktivierungsfunktionen modulieren. Diese sind jedoch lediglich externe Optimierungen, keine internen Systeme, die Aktivität und Plastizität in Echtzeit regulieren. Wie Mei, Müller & Ramaswamy (2022) in Trends in Neurosciences argumentierten, bleibt es eine offene Herausforderung, tiefe neuronale Netzwerke durch multiskalare Prinzipien von neuromodulatorischen Systemen zu informieren, eine Herausforderung, die die aktuellen LLM-Architekturen nicht angesprochen haben.
Obwohl Neuromodulation manchmal in KI-Kontexten erwähnt wird, bleiben LLMs und Transformer teilweise Annäherungen, keine Systeme, die mit dem Gehirn vergleichbar sind. Die Kluft zwischen statischen Matrixberechnungen und der dynamischen, zustandsabhängigen Regulierung, die in biologischen neuronalen Netzwerken zu finden ist, macht gerade die hirninspirierten KI-Architekturen wie Neuraxon zu einem notwendigen nächsten Schritt in Richtung adaptiver künstlicher Intelligenz.
3. Wie Neuraxon Neuromodulation berechnet: Hirninspirierte KI-Architektur
In Neuraxon ist die Berechnung ein Prozess, der sich in kontinuierlicher Zeit entfaltet. Der Code drückt ein System aus, das interne Zustände, s(t), aufrechterhält, die sich sogar in Abwesenheit klarer externer Stimuli entwickeln. Diese Zustände beeinflussen zukünftiges Verhalten und schaffen ein lebendes neuronales System, das immer aktiv ist, ein Konzept, das im Neuraxon-Forschungspapier im Detail behandelt wird.
Schnelle, langsame und Meta-Dynamik in neuronaler Berechnung
Neuraxon integriert explizit schnelle, langsame und Meta-Dynamiken und spiegelt die multiskalare temporale Architektur wider, die im biologischen Gehirn zu finden ist. Schnelle Dynamik regiert die sofortige Ausbreitung von Aktivität, analog zu schneller neuronaler Signalgebung über ionotrope Rezeptoren. Langsame Dynamik führt zu Akkumulation, Persistenz und Stabilisierung von Mustern, wodurch das System Informationen über den Moment hinaus speichern kann, ähnlich wie metabotrope Rezeptoren die neuronale Funktion über Sekunden und Minuten modulieren. Meta-Dynamik wirkt auf die Regeln der Interaktion zwischen den vorherigen und moduliert, wann das System empfindlicher für Veränderungen wird und wann es dazu neigt, seinen Zustand zu bewahren.
Die Neuromodulation in Neuraxon wird nicht als externe Parameteranpassung implementiert. Das System entscheidet nicht explizit, was es lernen soll, sondern vielmehr unter welchen Bedingungen es sich ändern kann. Dies spiegelt wider, wie biologische Neuromodulatoren wie Dopamin und Serotonin Fenster der Plastizität schaffen, anstatt Informationen direkt zu kodieren. Sie können diese Dynamiken aus erster Hand mit der interaktiven 3D-Simulation von Neuraxon auf HuggingFace Spaces erkunden, wo Sie Dopamin-, Serotonin-, Acetylcholin- und Noradrenalinspiegel in Echtzeit anpassen und beobachten können, wie sie das Netzwerkverhalten beeinflussen.
Von biologischen Prinzipien zu dezentraler KI
Dieser Ansatz reproduziert nicht die molekulare oder anatomische Komplexität des Gehirns, was derzeit unmöglich zu replizieren ist. Es gibt keine Tausenden von Rezeptoren oder echten biologischen Netzwerken. Es bewahrt jedoch ein essentielles Prinzip: Intelligenz ist adaptiv und erfordert daher interne Dynamik, Zustand und Modulation.
Die Neuromodulationsarchitektur von Neuraxon ist ein zentraler Bestandteil von Qubics umfassender Vision für dezentrale KI. Durch die Integration von Neuraxon mit dem Aigarth Intelligent Tissue-evolutionären Rahmen schafft Qubic ein System, in dem Millionen von auf Neuraxon basierenden Architekturen sich entwickeln, konkurrieren und durch verteilte Berechnung verbessern können, unterstützt durch den Konsensmechanismus des Qubic-Netzwerks, den nützlichen Proof of Work (UPoW).
4. Erforschen Sie Neuromodulatoren mit der interaktiven Neuraxon-Demo
Möchten Sie erfahren, wie Neuromodulation in einem hirninspirierten KI-System funktioniert? Die Neuraxon Mood Mixer-Demo ermöglicht es Ihnen, Dopamin-, Serotonin-, Acetylcholin- und Noradrenalinspiegel in Echtzeit anzupassen und zu beobachten, wie diese Neuromodulatoren das Verhalten neuronaler Netzwerke beeinflussen. Es ist eine praktische Möglichkeit, die in diesem Artikel diskutierten Prinzipien zu verstehen und den Unterschied zwischen statischer KI-Berechnung und dynamischer, zustandsabhängiger Verarbeitung zu sehen.
5. Die Mathematik hinter der multiskalaren Neuromodulation von Neuraxon
Die zeitlichen Dynamiken in Neuraxon werden von drei Differentialgleichungen geregelt, die die schnellen, langsamen und meta Zeitskalen neuronaler Berechnung erfassen:

Hier spiegeln τ_fast < τ_slow < τ_meta ihre unterschiedlichen zeitlichen Skalen wider, wobei τ_meta signifikant größer ist, um die 'ultraschnelle' Natur der metabotropen Effekte zu erfassen. Dieses mathematische Framework implementiert direkt das biologische Prinzip, dass Neuromodulation auf viel langsameren Zeitskalen als schnelle synaptische Übertragung wirkt, wie von Northoff & Huang (2017) in ihrer Arbeit beschrieben, wie die zeitlichen Dynamiken des Gehirns das Bewusstsein vermitteln.
Wissenschaftliche Referenzen
Dayan, P., & Huys, Q. J. M. (2009). Serotonin, Hemmung und negative Stimmung. PLoS Computational Biology.
Marder, E. (2012). Neuromodulation neuronaler Schaltkreise: zurück in die Zukunft. Neuron.
Schultz, W. (2016). Dopamin-Belohnungsprognosefehlerkodierung. Dialoge in klinischer Neurowissenschaft.
Aston-Jones, G., & Cohen, J. D. (2005). Eine integrative Theorie der Funktion von locus coeruleus–Norepinephrin. Jahresüberblick der Neurowissenschaft.
Mei, L., Müller, E., & Ramaswamy, S. (2022). Tiefe neuronale Netzwerke durch multiskalare Prinzipien neuromodulatorischer Systeme informieren. Trends in Neurowissenschaften.
Friston, K. (2010). Das Prinzip der freien Energie: eine einheitliche Gehirntheorie? Nature Reviews Neuroscience.
Northoff, G., & Huang, Z. (2017). Wie vermitteln die Zeit- und Raumdynamiken des Gehirns Bewusstsein und seine Störungen? Bewusstsein und Kognition, 57, 1–10.