
Scris de echipa științifică QubicRaportul de ramificare și criticalitatea în rețelele biologice, în rețelele artificiale și ca principiu bioinspirat în Neuraxon
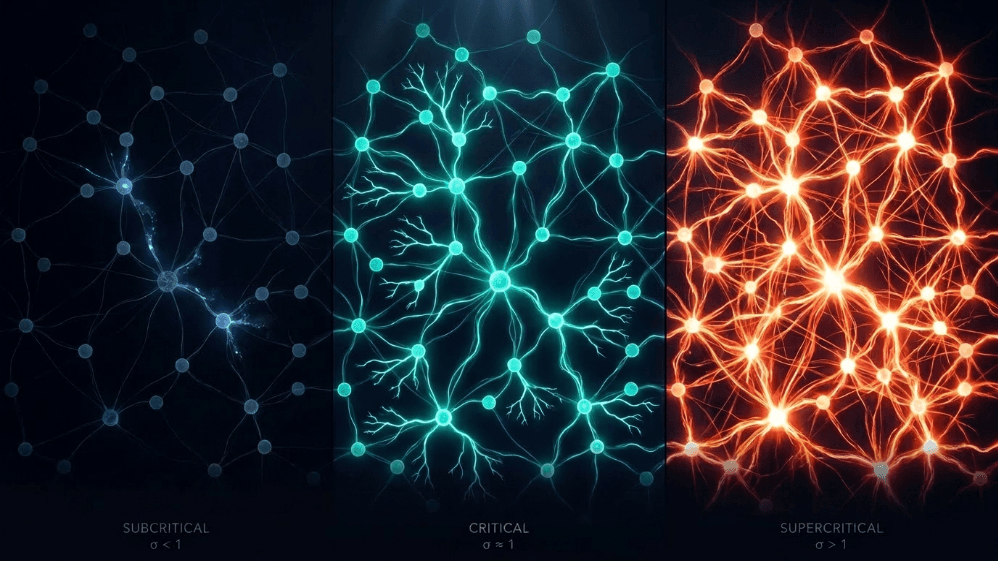
Ce au în comun o avalanșă de zăpadă, un incendiu de pădure, un cutremur și activitatea spontană a cortexului cerebral?
Toate au în comun o graniță între ordine și haos, ceea ce se numește o stare critică. În creier, această limită este măsurată printr-un parametru simplu: raportul de ramificare (σ sau m). Ar fi ceva similar cu raportul mediu al „descendenților” neuronali pe care fiecare neuron „părinte” îi activează. Când σ ≈ 1, activitatea nici nu se stinge, nici nu explodează; ea reverberează.
Beggs și Plenz (2003) au înregistrat activitatea spontană a cortexului cerebral la șobolani și au descoperit că activitatea a format modele asemănătoare cascadei, așa-numitele avalanșe neuronale, cu un raport de ramificare apropiat de 1. Creierul părea să trăiască într-un punct critic. La oameni, raportul de ramificare σ apare din nou aproape de unitate (Wang et al., 2025; Plenz et al., 2021; Wilting & Priesemann, 2019).
În punctul critic, sistemele prezintă simultan sensibilitate maximă la perturbații (capacitate de răspuns), capacitate dinamică maximă (numărul de stări accesibile), transmitere maximă de informații și complexitate maximă (Timme et al., 2016; Shew et al., 2009, 2011).
Ce este raportul de ramificare și cum se măsoară?
Conceptual, raportul de ramificare este banal: dacă la momentul t există A(t) neuroni activi și la t+1 există A(t+1), atunci:
σ = ⟨ A(t+1) / A(t) ⟩
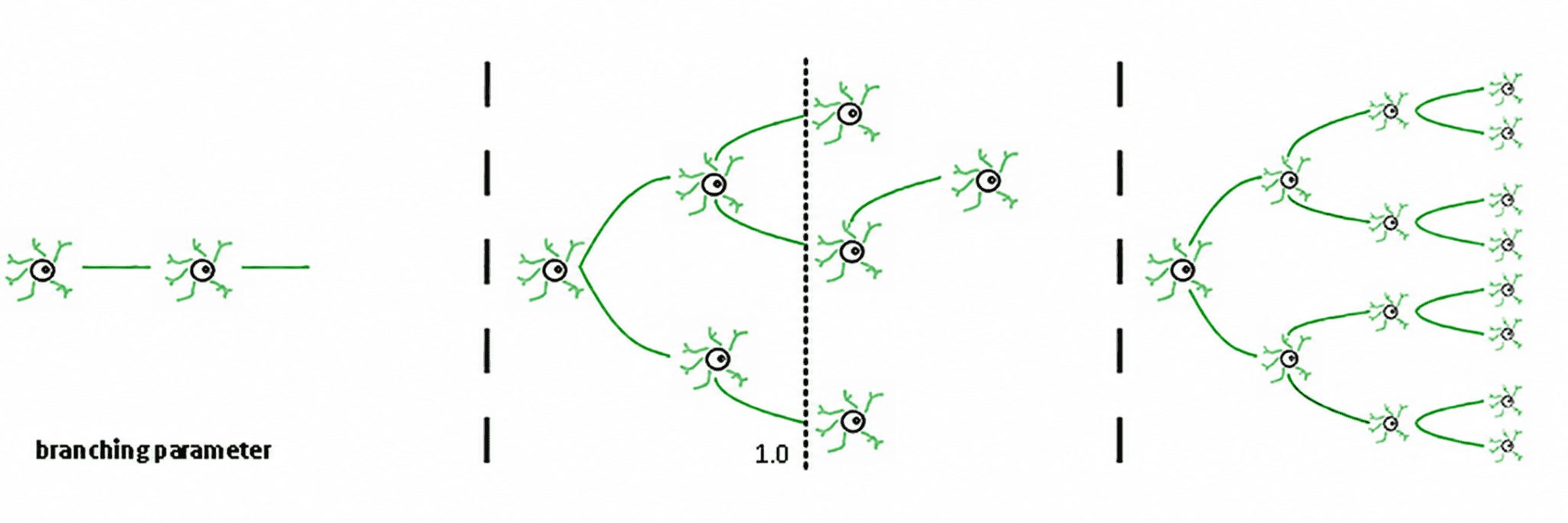
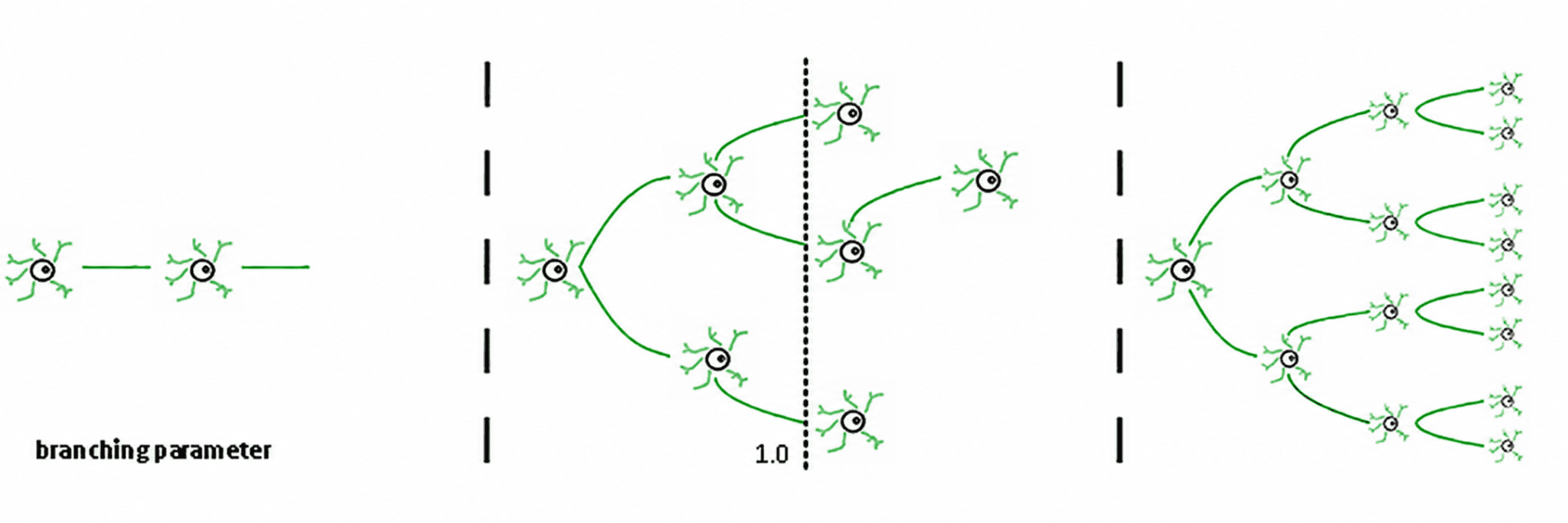
Din aceasta rezultă trei regimuri (de Carvalho și Prado, 2000; Haldeman și Beggs, 2005):
Subcritic (σ < 1): activitatea scade; sistemul „uită” rapid perturbația. Este stabil, dar are memorie slabă și nu este foarte expresiv.
Supercritic (σ > 1): activitatea explodează în cascade. Aceasta este semnătura regimurilor patologice, cum ar fi crizele epileptice (Hsu et al., 2008; Hagemann et al., 2021).
Critic (σ ≈ 1): fiecare vârf, în medie, generează un alt vârf. Activitatea reverberează, avalanșele neuronale respectă legile puterii, iar sistemul menține o memorie structurată a inputului.
Frumusețea lui σ constă în faptul că este un singur număr care rezumă regimul dinamic global. Însă măsurarea sa este mai puțin banală. Atunci când este aplicată înregistrărilor corticale in vivo, măsurarea dezvăluie că cortexul nu funcționează exact la σ = 1, ci puțin sub, într-un regim pe care autorii îl numesc reverberativ (Wilting et al., 2018). Diferența este importantă: a fi exact la σ = 1 ar fi ca și cum ai pedala o bicicletă echilibrată pe o frânghie întinsă; a fi puțin sub permite o adaptare rapidă la cerințele sarcinii fără riscul unei explozii fulgerătoare.
Criticitatea în rețelele neuronale artificiale: de la marginea haosului la calculul rezervorului
Bertschinger și Natschläger (2004) au arătat că rețelele cu prag recurent aleatoriu își ating capacitatea maximă de calcul în sarcinile de procesare temporală exact la tranziția ordine-haos.
Boedecker și colab. (2012) au extins analiza la rețelele de stări ecou din cadrul paradigmei de calcul rezervor, confirmând că capacitatea de transfer de informații și memoria activă sunt maximizate la limita haosului.
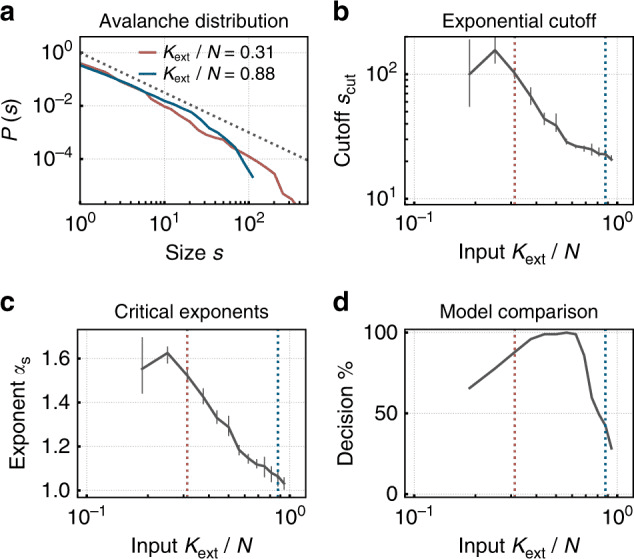
Fig. 3. O rețea neuromorfă cu spiking și plasticitate sinaptică se auto-organizează spre critică în condiții de input extern scăzut, prezentând distribuții ale dimensiunii avalanșelor de tip lege de putere - semnul distinctiv al stării critice atât în rețelele neuronale biologice, cât și în cele artificiale. În condiții de input mai mare, rețeaua trece la un regim subcritic cu distribuții trunchiate. Reprodus din Cramer și colab. (2020), Nature Communications, 11, 2853. CC BY 4.0.
În limbajul rețelelor neuronale artificiale, parametrul de măsurare se numește rază spectrală. Când depășește 1, traiectoriile diverg exponențial (haos); când este mult sub 1, rețeaua se prăbușește până la punctul fix și își pierde memoria. Raza spectrală apropiată de 1 este, în acest context, echivalentul formal al valorii biologice σ ≈ 1 (Magnasco, 2022; Morales et al., 2023). În rețelele neuronale de tip spiking, raportul de ramificare poate fi măsurat cu metode aproape identice cu cele utilizate în culturile neuronale (Cramer et al., 2020; Zeraati et al., 2024).
De ce maximizează criticitatea creierului calculul neuronal?
Funcționarea aproape de σ ≈ 1 oferă patru avantaje care sunt esențiale atât pentru ipoteza creierului critic, cât și pentru proiectarea sistemelor de inteligență artificială inspirate de creier:
Interval dinamic maxim. Shew și colab. (2009) au arătat că intervalul de intensități de intrare pe care cortexul le poate discrimina este maxim atunci când echilibrul excitație-inhibiție plasează rețeaua la nivel critic.
Capacitate informațională maximizată. Entropia modelelor de avalanșă și informația reciprocă dintre vârful de intrare și cel de ieșire la σ ≈ 1 (Shew et al., 2011).
Memorie optimă în decolorare. În regimul critic, perturbația este susținută suficient de mult timp pentru a influența procesarea fără a contamina viitorul îndepărtat; este punctul ideal dintre stabilitate și integrare temporală (Boedecker et al., 2012).
Complexitatea ca măsură unificatoare. Timme și colab. (2016) au demonstrat că complexitatea neuronală este maximizată exact în punctul critic, corelând criticitatea cu teoriile formale ale conștiinței și procesării.
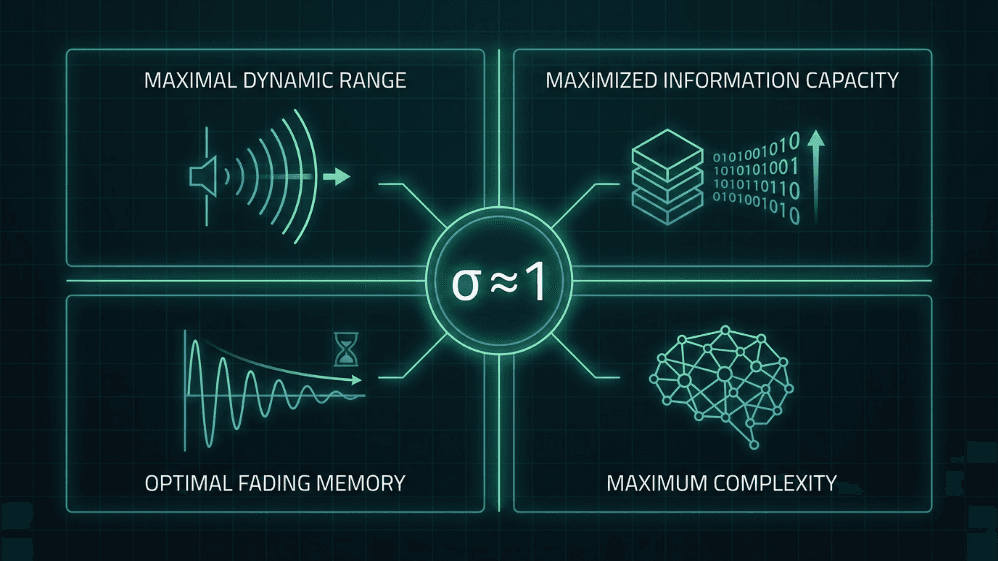
Fig. 4. Patru avantaje computaționale ale funcționării în apropierea raportului critic de ramificare (σ ≈ 1). La nivel critic, rețelele neuronale ating o gamă dinamică maximă, o capacitate informațională maximă, o memorie optimă de fading și o complexitate maximă - proprietăți care sunt esențiale atât pentru ipoteza creierului critic, cât și pentru designul IA inspirat de creier.
Creierul nu funcționează întotdeauna la σ = 1
Aceasta nu implică faptul că creierul funcționează întotdeauna la σ = 1. Dovezile sugerează mai degrabă un regim ușor subcritic și modulabil: în timpul sarcinilor solicitante, rețeaua se apropie de criticitate, în timpul somnului profund se îndepărtează, iar stările patologice (epilepsie, anestezie profundă, anumite afecțiuni psihiatrice) sunt asociate cu abateri măsurabile de la acest interval operațional (Meisel et al., 2017; Zimmern, 2020). Raportul de ramificare devine un biomarker dinamic al stării funcționale a sistemului nervos.
De ce folosim raportul de ramificare în Neuraxon: Design AI bioinspirat la marginea haosului
Neuraxon este un sistem bioinspirat care adoptă principii dinamice ale cortexului ca și constrângeri de design. Raportul de ramificare este unul dintre cele mai importante și îl folosim din patru motive:
Ca invariant operațional în timp real pentru stabilitatea rețelei neuronale
În arhitecturile recurente sau cu vârfuri mari de activitate, riscul dual de colaps al activității (rețea silențioasă, gradienți care dispar) și explozie fugătoare (saturație, gradienți care explodează) este structural. Monitorizarea lui σ în timp real ne oferă un singur scalar de diagnostic, independent de arhitectura concretă, care indică dacă sistemul este activ în sens computațional.
Ca o țintă de autoreglare bioinspirată prin criticitate autoorganizată
Rețeaua se auto-organizează către o anumită importanță critică fără a fi nevoie de o ajustare centralizată, replicând principiul auto-organizării critice (Bornholdt & Röhl, 2003; Levina et al., 2007). Acest lucru reduce drastic sensibilitatea la hiperparametri și conferă sistemului robustețe împotriva schimbărilor de distribuție. Așa cum am explorat în NIA Volumul 7 despre viața artificială și ecosistemele digitale, exact așa apare complexitatea emergentă din regulile locale fără control centralizat.
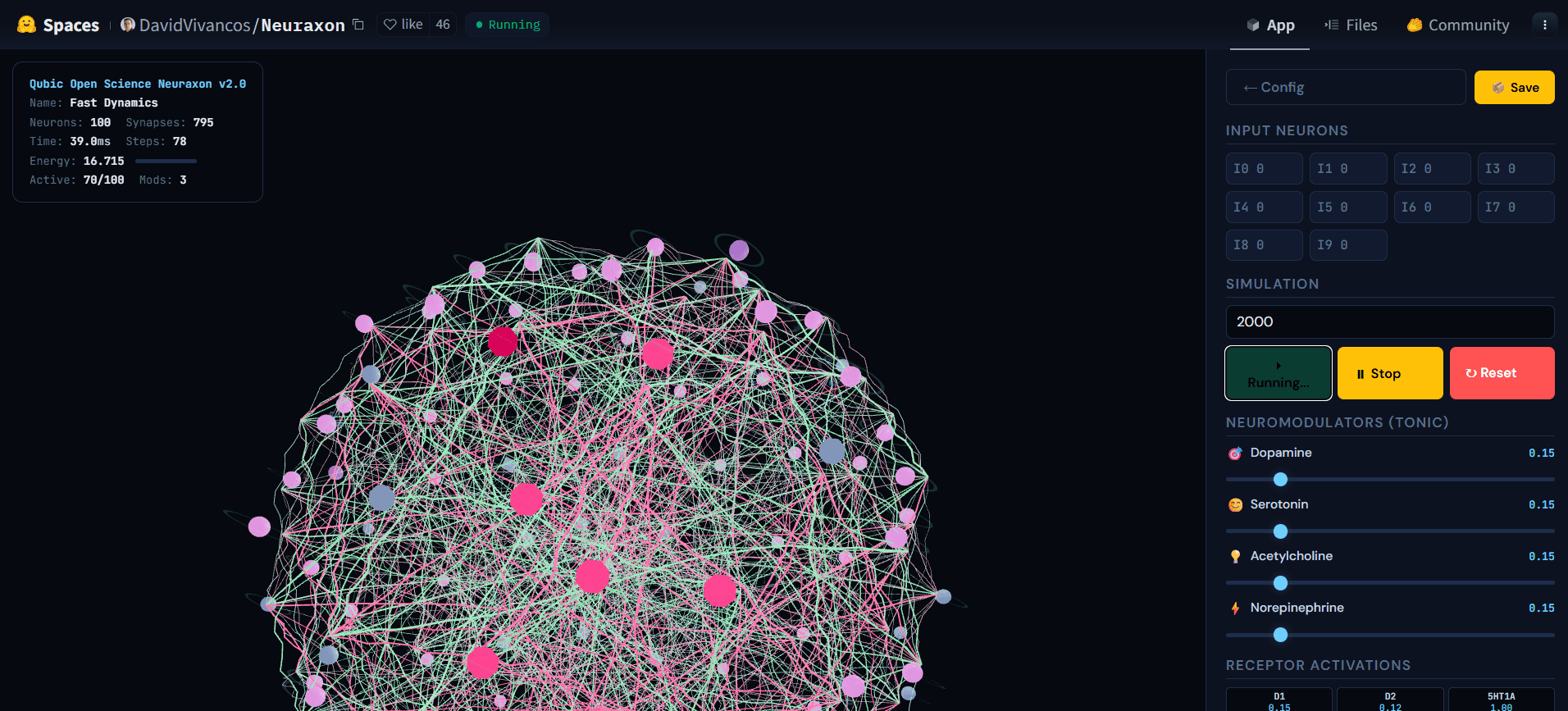
Fig. 5. Rețeaua 3D Neuraxon în timpul simulării active, arătând activitatea în cascadă între neuronii în stare ternară. Nodurile puternic active (roz) propagă semnale prin conexiuni excitatorii (verde) și inhibitorii (roz), în timp ce alți neuroni rămân în repaus (gri), ilustrând un regim reverberant în apropierea raportului critic de ramificare (σ ≈ 1). Această stare echilibrată - nici silențioasă, nici explozivă - este ceea ce Neuraxon auto-organizează folosind principii de criticitate bioinspirate. Explorați demonstrația interactivă athuggingface.co/spaces/DavidVivancos/Neuraxon. Sursa: Echipa științifică Qubic.
Ca o punte între observația neuroștiințifică și designul inteligenței artificiale
Raportul de ramificare este una dintre puținele magnitudini măsurate cu același formalism în electrofiziologie, RMN funcțional și rețele artificiale. Acest lucru permite testarea ipotezelor bidirecționale: dacă o intervenție îmbunătățește criticitatea biologică, ne putem întreba dacă aceeași intervenție - tradusă în arhitectura artificială - îmbunătățește calculul modelului și invers. Acest principiu este esențial pentru cadrul de neuromodulație și mecanismele de sincronizare astrocitară pe care le-am dezvoltat în volumele anterioare ale acestei academii.
Ca criteriu funcțional, nu estetic, pentru IA inspirată de creier
Criticitatea este o constrângere operațională cu consecințe empirice. Operarea în apropierea regimului reverberant îmbunătățește — așa cum este măsurat în evaluările noastre interne și în publicațiile trimise — capacitatea de generalizare, stabilitatea în fața perturbațiilor de input, bogăția reprezentațională și coerența temporală a raționamentului. Aceste efecte corespund calitativ cu cele raportate atât în literatura biologică (Cocchi et al., 2017), cât și în cea artificială (Cramer et al., 2020; Morales et al., 2023).
Raportul de ramificare: de la fizica statistică la arhitectura IA inspirată de creier
Raportul de ramificare este una dintre acele rarități conceptuale: suficient de simplu pentru a fi redus la o singură formulă, suficient de profund pentru a face legătura între fizica statistică, neuroștiință, inteligența artificială și proiectarea sistemelor. Pentru creierul biologic, σ ≈ 1 pare a fi regimul în care apare combinația virtuoasă de sensibilitate, memorie, expresivitate și robustețe. Pentru rețelele artificiale, aceeași frontieră - redenumită marginea haosului - prezice capacitatea computațională maximă.
Și pentru Neuraxon, este un principiu călăuzitor al designului bioinspirat: o metrică auditabilă, autoreglatoare și biologic semnificativă, care ne ajută să menținem sistemul în viață, în cel mai bogat sens al cuvântului.
Referințe
Beggs, J. M. și Plenz, D. (2003). Avalanșe neuronale în circuitele neocorticale. The Journal of Neuroscience, 23(35), 11167–11177. https://doi.org/10.1523/JNEUROSCI.23-35-11167.2003
Bertschinger, N. și Natschläger, T. (2004). Calcul în timp real la limita haosului în rețelele neuronale recurente. Neural Computation, 16(7), 1413–1436. https://doi.org/10.1162/089976604323057443
Boedecker, J., Obst, O., Lizier, J. T., Mayer, N. M. și Asada, M. (2012). Prelucrarea informațiilor în rețelele de stări ecou la marginea haosului. Theory in Biosciences, 131(3), 205–213. https://doi.org/10.1007/s12064-011-0146-8
Bornholdt, S. și Röhl, T. (2003). Rețele neuronale critice auto-organizate. Physical Review E, 67(6), 066118. https://doi.org/10.1103/PhysRevE.67.066118
Cocchi, L., Gollo, L. L., Zalesky, A. și Breakspear, M. (2017). Importanța creierului: o sinteză a neurobiologiei, modelelor și cunoașterii. Progress in Neurobiology, 158, 132–152. https://doi.org/10.1016/j.pneurobio.2017.07.002
Cramer, B., Stöckel, D., Kreft, M., Wibral, M., Schemmel, J., Meier, K. și Priesemann, V. (2020). Controlul criticității și al calculului în adăugarea de plasticitate rețelelor neuromorfice. Nature Communications, 11, 2853. https://doi.org/10.1038/s41467-020-16548-3
de Carvalho, J. X. și Prado, C. P. C. (2000). Criticitate auto-organizată în modelul Olami-Feder-Christensen. Physical Review Letters, 84(17), 4006–4009. https://doi.org/10.1103/PhysRevLett.84.4006
Derrida, B. și Pomeau, Y. (1986). Rețele aleatoare de automate: o aproximare simplă prin recoacere. Europhysics Letters, 1(2), 45–49. https://doi.org/10.1209/0295-5075/1/2/001
Hagemann, A., Wilting, J., Samimizad, B., Mormann, F. și Priesemann, V. (2021). Evaluarea criticității activității neuronale individuale din cortexul epileptic uman, înainte de criză. PLOS Computational Biology, 17(3), e1008773. https://doi.org/10.1371/journal.pcbi.1008773
Haldeman, C. și Beggs, J. M. (2005). Ramificarea critică surprinde activitatea în rețelele neuronale vii și maximizează numărul de stări metastabile. Physical Review Letters, 94(5), 058101. https://doi.org/10.1103/PhysRevLett.94.058101
Hsu, D., Chen, W., Hsu, M. și Beggs, J. M. (2008). O ipoteză deschisă: Epilepsia este învățată și poate fi dezvățată? Epilepsy & Behavior, 13(3), 511–522. https://doi.org/10.1016/j.yebeh.2008.05.007
Langton, C. G. (1990). Calcul la marginea haosului: Tranziții de fază și calcul emergent. Physica D: Nonlinear Phenomena, 42(1–3), 12–37. https://doi.org/10.1016/0167-2789(90)90064-V
Levina, A., Herrmann, J. M. și Geisel, T. (2007). Sinapse dinamice care cauzează criticitate auto-organizată în rețelele neuronale. Nature Physics, 3(12), 857–860. https://doi.org/10.1038/nphys758
Magnasco, M. O. (2022). Robustețea și flexibilitatea funcției neuronale prin criticitate dinamică. Entropy, 24(5), 591. https://doi.org/10.3390/e24050591
Meisel, C., Klaus, A., Vyazovskiy, V. V. și Plenz, D. (2017). Interacțiunea dintre corelațiile temporale pe termen lung și scurt modelează dinamica cortexului în stările de vigilență. The Journal of Neuroscience, 37(42), 10114–10124. https://doi.org/10.1523/JNEUROSCI.0448-17.2017
Morales, G. B., di Santo, S. și Muñoz, M. A. (2023). Dezvăluirea dinamicii intrinseci a rețelelor neuronale biologice și artificiale: De la criticitate la reprezentări optime. Frontiers in Complex Systems, 1, 1276338. https://doi.org/10.3389/fcpxs.2023.1276338
Plenz, D., Ribeiro, T. L., Miller, S. R., Kells, P. A., Vakili, A. și Capek, E. L. (2021). Criticitate auto-organizată în creier. Frontiers in Physics, 9, 639389. https://doi.org/10.3389/fphy.2021.639389
Shew, W. L., Yang, H., Petermann, T., Roy, R. și Plenz, D. (2009). Avalanșele neuronale implică o gamă dinamică maximă în rețelele corticale la nivel critic. The Journal of Neuroscience, 29(49), 15595–15600. https://doi.org/10.1523/JNEUROSCI.3864-09.2009
Shew, W. L., Yang, H., Yu, S., Roy, R. și Plenz, D. (2011). Capacitatea și transmiterea informației sunt maximizate în rețelele corticale echilibrate cu avalanșe neuronale. The Journal of Neuroscience, 31(1), 55–63. https://doi.org/10.1523/JNEUROSCI.4637-10.2011
Spitzner, F. P., Dehning, J., Wilting, J., Hagemann, A., Neto, J. P., Zierenberg, J. și Priesemann, V. (2021). MR. Estimator, un set de instrumente pentru determinarea intervalelor de timp intrinseci din activitatea de spiking subeșantionată. PLOS ONE, 16(4), e0249447. https://doi.org/10.1371/journal.pone.0249447
Timme, N. M., Marshall, N. J., Bennett, N., Ripp, M., Lautzenhiser, E. și Beggs, J. M. (2016). Criticitatea maximizează complexitatea în țesutul neural. Frontiers in Physiology, 7, 425. https://doi.org/10.3389/fphys.2016.00425
Turrigiano, G. G. (2008). Neuronul autoreglabil: Scalarea sinaptică a sinapselor excitatorii. Cell, 135(3), 422–435. https://doi.org/10.1016/j.cell.2008.10.008
Wang, J., Cao, R., Brunton, B. W., Smith, R. E. W., Buckner, R. L. și Liu, T. T. (2025). Contribuții genetice la capacitatea critică a creierului și relația acesteia cu funcțiile cognitive umane. Proceedings of the National Academy of Sciences, 122(26), e2417010122. https://doi.org/10.1073/pnas.2417010122
Wilting, J., Dehning, J., Pinheiro Neto, J., Rudelt, L., Wibral, M., Zierenberg, J. și Priesemann, V. (2018). Funcționarea într-un regim reverberant permite ajustarea rapidă a stărilor rețelei la cerințele sarcinii. Frontiers in Systems Neuroscience, 12, 55. https://doi.org/10.3389/fnsys.2018.00055
Wilting, J. și Priesemann, V. (2018). Deducerea stărilor dinamice colective din sisteme neobservate în mare parte. Nature Communications, 9, 2325. https://doi.org/10.1038/s41467-018-04725-4
Wilting, J. și Priesemann, V. (2019). 25 de ani de criticitate în neuroștiințe — Rezultate stabilite, controverse deschise, concepte noi. Current Opinion in Neurobiology, 58, 105–111. https://doi.org/10.1016/j.conb.2019.08.002
Yu, C. (2022). Către o analiză unificată a ipotezei criticității creierului: Revizuirea mai multor instrumente disponibile. Frontiers in Neural Circuits, 16, 911245. https://doi.org/10.3389/fncir.2022.911245
Zeraati, R., Engel, T. A. și Levina, A. (2024). Estimarea intervalelor de timp intrinseci și a criticității din înregistrările neuronale: metode și capcane. Current Opinion in Neurobiology, 86, 102871. https://doi.org/10.1016/j.conb.2024.102871
Zimmern, V. (2020). De ce este relevantă clinic importanța creierului: o analiză a domeniului de aplicare. Frontiers in Neural Circuits, 14, 54. https://doi.org/10.3389/fncir.2020.00054
Explorează Academia Completă de Inteligență Neuraxon
Acesta este volumul 8 din cartea #Neuraxon Intelligence #academy realizată de echipa științifică #Qubic . Dacă abia vă alăturați nouă, explorați seria completă pentru a înțelege pe deplin știința din spatele Neuraxon, #aigarth și abordarea lui Qubic asupra inteligenței artificiale inspirate de creier, #decentralized :
NIA Vol. 1: De ce inteligența nu este calculată în etape, ci în timp — Explorează de ce inteligența biologică funcționează în timp continuu, mai degrabă decât în etape computaționale discrete, cum ar fi LLM-urile tradiționale.
NIA Vol. 2: Dinamica ternară ca model al inteligenței vii — Explică dinamica ternară și de ce logica în trei stări (excitatorie, neutră, inhibitorie) este importantă pentru modelarea sistemelor vii.
NIA Vol. 3: Neuromodulare și inteligență artificială inspirată de creier — Acoperă neuromodularea și modul în care semnalizarea chimică a creierului (dopamină, serotonină, acetilcolină, norepinefrină) inspiră arhitectura Neuraxon.
NIA Vol. 4: Rețele neuronale în inteligență artificială și neuroștiințe — O comparație aprofundată între rețelele neuronale biologice, rețelele neuronale artificiale și abordarea cu a treia cale a Neuraxon.
NIA Vol. 5: Astrocite și inteligență artificială inspirată de creier — Cum transformă sincronizarea astrocitară plasticitatea rețelei neuronale prin intermediul cadrului AGMP din Neuraxon.
NIA Vol. 6: Mașini conștiente vs. organisme inteligente: Conștiința IA explicată — Explorează conștiința IA prin prisma teoriei spațiului de lucru global, a teoriei informației integrate și a codării predictive.
NIA Vol. 7: Jocul vieții, viața artificială și ecosistemele digitale al lui Conway — Știința din spatele abordării lui Qubic, Aigarth și Neuraxon asupra complexității emergente și a criticității auto-organizate în IA descentralizată.
Qubic este o rețea descentralizată, open-source, pentru tehnologie experimentală. Pentru a afla mai multe, vizitați qubic.org. Alăturați-vă discuției pe X, Discord și Telegram.