
Ditulis oleh Tim Ilmiah Qubic. Rasio percabangan dan kritisitas dalam jaringan biologis, dalam jaringan buatan, dan sebagai prinsip terinspirasi biologis dalam Neuraxon.
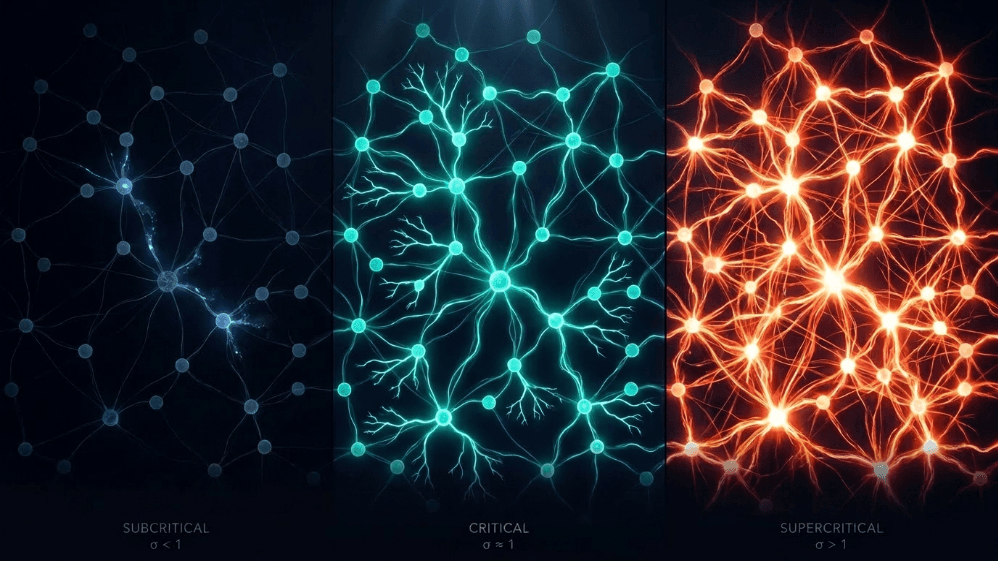
Apa kesamaan antara longsoran salju, kebakaran hutan, gempa bumi, dan aktivitas spontan korteks serebral?
Mereka semua berbagi batas antara order dan kekacauan, yang disebut sebagai keadaan kritis. Di otak, tepi itu diukur dengan parameter sederhana: rasio percabangan (σ atau m). Ini akan menjadi semacam rasio rata-rata dari 'anak' neuronal yang diaktifkan oleh setiap neuron 'induk'. Ketika σ ≈ 1, aktivitas tidak mati atau meledak; itu bergetar.
Beggs dan Plenz (2003) merekam aktivitas spontan korteks serebral pada tikus dan menemukan bahwa aktivitas membentuk pola mirip longsor, yang disebut longsor neuronal, dengan rasio cabang mendekati 1. Otak tampaknya hidup di titik kritis. Pada manusia, rasio cabang σ sekali lagi muncul mendekati satu (Wang et al., 2025; Plenz et al., 2021; Wilting & Priesemann, 2019).
Pada titik kritis, sistem secara bersamaan menunjukkan sensitivitas maksimal terhadap gangguan (responsivitas), kapasitas dinamis maksimum (jumlah status yang dapat diakses), transmisi informasi maksimum, dan kompleksitas maksimum (Timme et al., 2016; Shew et al., 2009, 2011).
Apa itu Rasio Cabang dan Bagaimana Cara Mengukurnya?
Secara konseptual, rasio cabang adalah sepele: jika pada saat t ada A(t) neuron aktif dan pada t+1 ada A(t+1), maka:
σ = ⟨ A(t+1) / A(t) ⟩
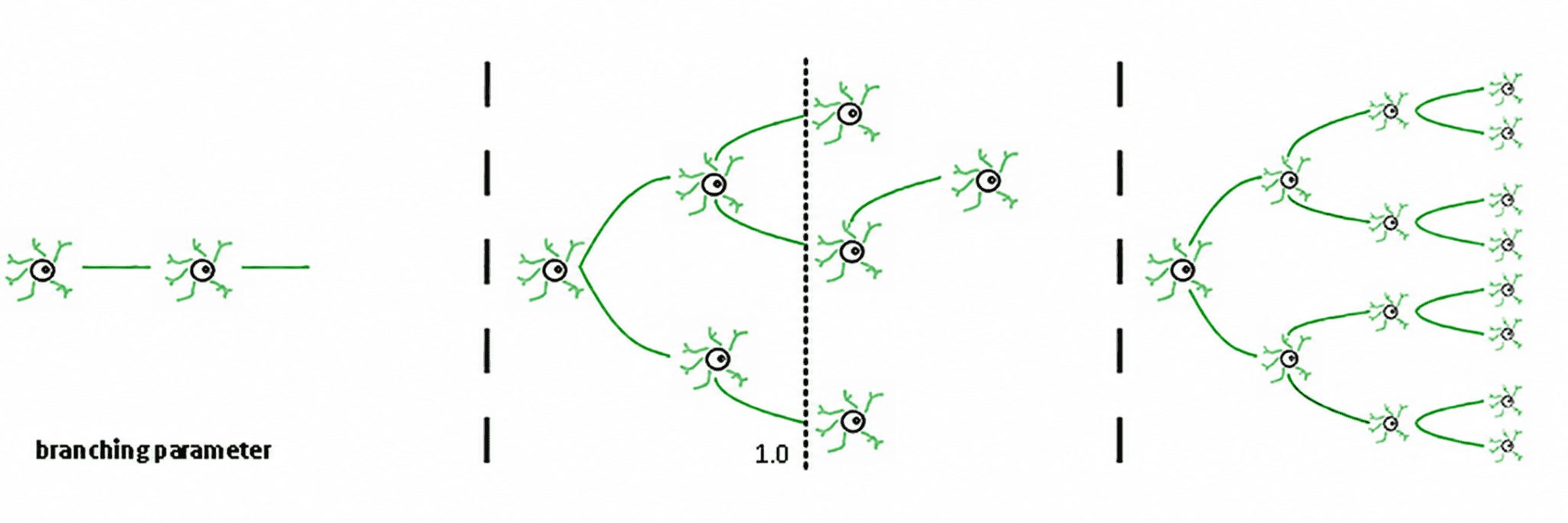
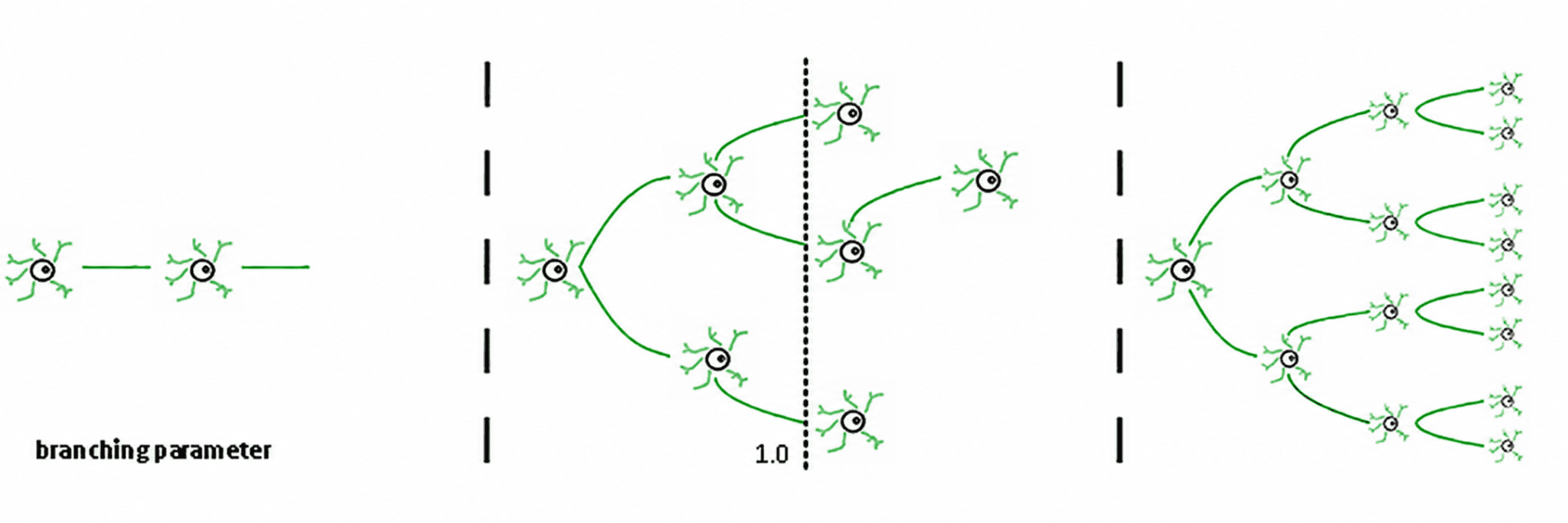
Tiga rezim mengikuti dari ini (de Carvalho & Prado, 2000; Haldeman & Beggs, 2005):
Subkritikal (σ < 1): aktivitas memudar; sistem "melupakan" gangguan dengan cepat. Ini stabil tetapi miskin dalam memori dan tidak terlalu ekspresif.
Superkritikal (σ > 1): aktivitas meledak menjadi cascades. Ini adalah ciri dari rezim patologis seperti kejang epilepsi (Hsu et al., 2008; Hagemann et al., 2021).
Kritis (σ ≈ 1): setiap spike, rata-rata, menghasilkan spike lain. Aktivitas bergetar, longsor neuronal mematuhi hukum kekuasaan, dan sistem mempertahankan memori terstruktur dari input.
Keindahan σ adalah bahwa ia adalah satu angka yang merangkum rezim dinamis global. Namun, mengukurnya kurang trivial. Ketika diterapkan pada rekaman kortikal in vivo, pengukuran mengungkapkan bahwa korteks tidak beroperasi persis pada σ = 1, tetapi sedikit di bawah, dalam rezim yang oleh penulis disebut reverberating (Wilting et al., 2018). Perbedaan ini penting: berada tepat di σ = 1 akan seperti mengayuh sepeda yang seimbang di atas tali; sedikit di bawah memungkinkan penyesuaian cepat terhadap tuntutan tugas tanpa risiko ledakan yang tak terkendali.
Kritisitas dalam Jaringan Neural Buatan: Dari Tepi Kekacauan ke Komputasi Reservoir
Bertschinger dan Natschläger (2004) menunjukkan bahwa jaringan ambang acak berulang mencapai kapasitas komputasi maksimumnya pada tugas pemrosesan temporal tepat di transisi urutan-kekacauan.
Boedecker et al. (2012) memperluas analisis ke jaringan echo state dalam paradigma komputasi reservoir, mengonfirmasi bahwa kapasitas transfer informasi dan memori aktif dimaksimalkan di tepi kekacauan.
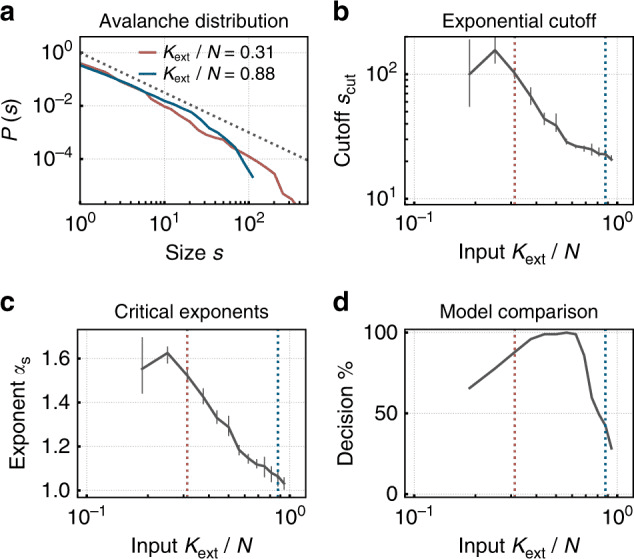
Gambar 3. Jaringan neuromorfik spiking dengan plastisitas sinaptik terorganisir sendiri menuju kritisitas di bawah input eksternal rendah, menunjukkan distribusi ukuran longsor hukum kekuasaan — ciri khas dari keadaan kritis baik dalam jaringan neural biologis maupun buatan. Di bawah input yang lebih tinggi, jaringan beralih ke rezim subkritikal dengan distribusi terputus. Diambil dari Cramer et al. (2020), Nature Communications, 11, 2853. CC BY 4.0.
Dalam bahasa jaringan neural buatan, parameter pengukuran disebut radius spektral. Ketika melebihi 1, lintasan menyimpang secara eksponensial (kekacauan); ketika jauh di bawah 1, jaringan runtuh ke titik tetap dan kehilangan memori. Radius spektral mendekati 1 adalah, dalam konteks ini, setara formal dari biologis σ ≈ 1 (Magnasco, 2022; Morales et al., 2023). Dalam jaringan neural spiking, rasio cabang dapat diukur dengan metode yang hampir identik dengan yang digunakan dalam kultur neuronal (Cramer et al., 2020; Zeraati et al., 2024).
Mengapa Kritisitas Otak Memaksimalkan Komputasi Neural?
Beroperasi dekat dengan σ ≈ 1 memberikan empat keuntungan yang penting untuk hipotesis otak kritis dan desain sistem AI terinspirasi otak:
Rentang dinamis maksimal. Shew et al. (2009) menunjukkan bahwa rentang intensitas input yang dapat dibedakan oleh korteks adalah maksimal ketika keseimbangan eksitasi-inhibisi menempatkan jaringan pada kritisitas.
Kapasitas informasi dimaksimalkan. Entropi pola longsor dan informasi mutual antara input dan output memuncak pada σ ≈ 1 (Shew et al., 2011).
Memori memudar optimal. Dalam rezim kritis, gangguan dipertahankan cukup lama untuk mempengaruhi pemrosesan tanpa mencemari masa depan yang jauh; ini adalah titik manis antara stabilitas dan integrasi temporal (Boedecker et al., 2012).
Kompleksitas sebagai ukuran penyatu. Timme et al. (2016) menunjukkan bahwa kompleksitas neural dimaksimalkan tepat di titik kritis, mengaitkan kritisitas dengan teori formal kesadaran dan pemrosesan.
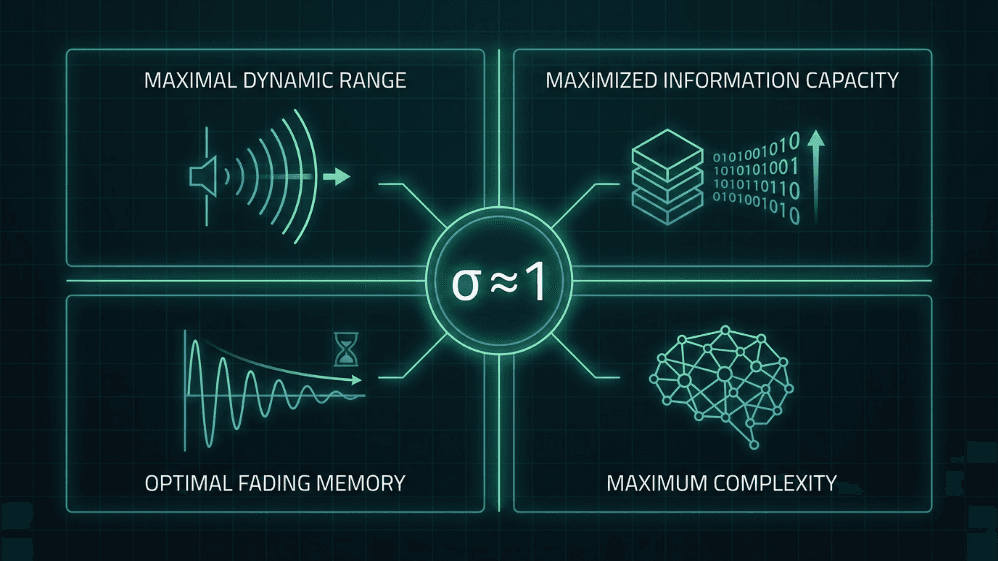
Gambar 4. Empat keuntungan komputasi dari beroperasi dekat dengan rasio cabang kritis (σ ≈ 1). Pada kritisitas, jaringan neural mencapai rentang dinamis maksimal, kapasitas informasi dimaksimalkan, memori memudar optimal, dan kompleksitas maksimum — properti yang menjadi pusat baik hipotesis otak kritis dan desain AI terinspirasi otak.
Otak Tidak Selalu Beroperasi pada σ = 1
Ini tidak menyiratkan bahwa otak selalu beroperasi pada σ = 1. Bukti lebih menunjukkan rezim yang sedikit subkritikal dan dapat dimodulasi: selama tugas yang menuntut jaringan mendekati kritisitas, selama tidur dalam ia bergerak menjauh, dan kondisi patologis (epilepsi, anestesi dalam, kondisi psikiatri tertentu) terkait dengan penyimpangan terukur dari rentang operasional ini (Meisel et al., 2017; Zimmern, 2020). Rasio cabang menjadi biomarker dinamis dari status fungsional sistem saraf.
Mengapa Kami Menggunakan Rasio Cabang di Neuraxon: Desain AI Terinspirasi Biologi di Tepi Kekacauan
Neuraxon adalah sistem terinspirasi biologi yang mengadopsi prinsip dinamis korteks sebagai batasan desain. Rasio cabang adalah salah satu yang paling penting, dan kami menggunakannya untuk empat alasan:
Sebagai Invarian Operasional Waktu Nyata untuk Stabilitas Jaringan Neural
Dalam arsitektur spiking atau berulang yang dalam, risiko ganda dari kolaps aktivitas (jaringan sunyi, gradien yang menghilang) dan ledakan tak terkendali (jenuh, gradien yang meledak) bersifat struktural. Memantau σ secara waktu nyata memberi kita satu skalar diagnostik, terlepas dari arsitektur konkret, yang menunjukkan apakah sistem aktif dalam arti komputasional.
Sebagai Target Regulasi Diri yang Terinspirasi Biologi Melalui Kritisitas Terorganisir Sendiri
Jaringan terorganisir sendiri menuju kritisitas tanpa perlu penyetelan halus terpusat, mereplikasi prinsip kritisitas terorganisir sendiri (Bornholdt & Röhl, 2003; Levina et al., 2007). Ini secara drastis mengurangi sensitivitas terhadap hiperparameter dan memberikan sistem ketahanan terhadap pergeseran distribusi. Seperti yang kami eksplorasi di NIA Volume 7 tentang kehidupan buatan dan ekosistem digital, inilah cara kompleksitas yang muncul muncul dari aturan lokal tanpa kontrol terpusat.
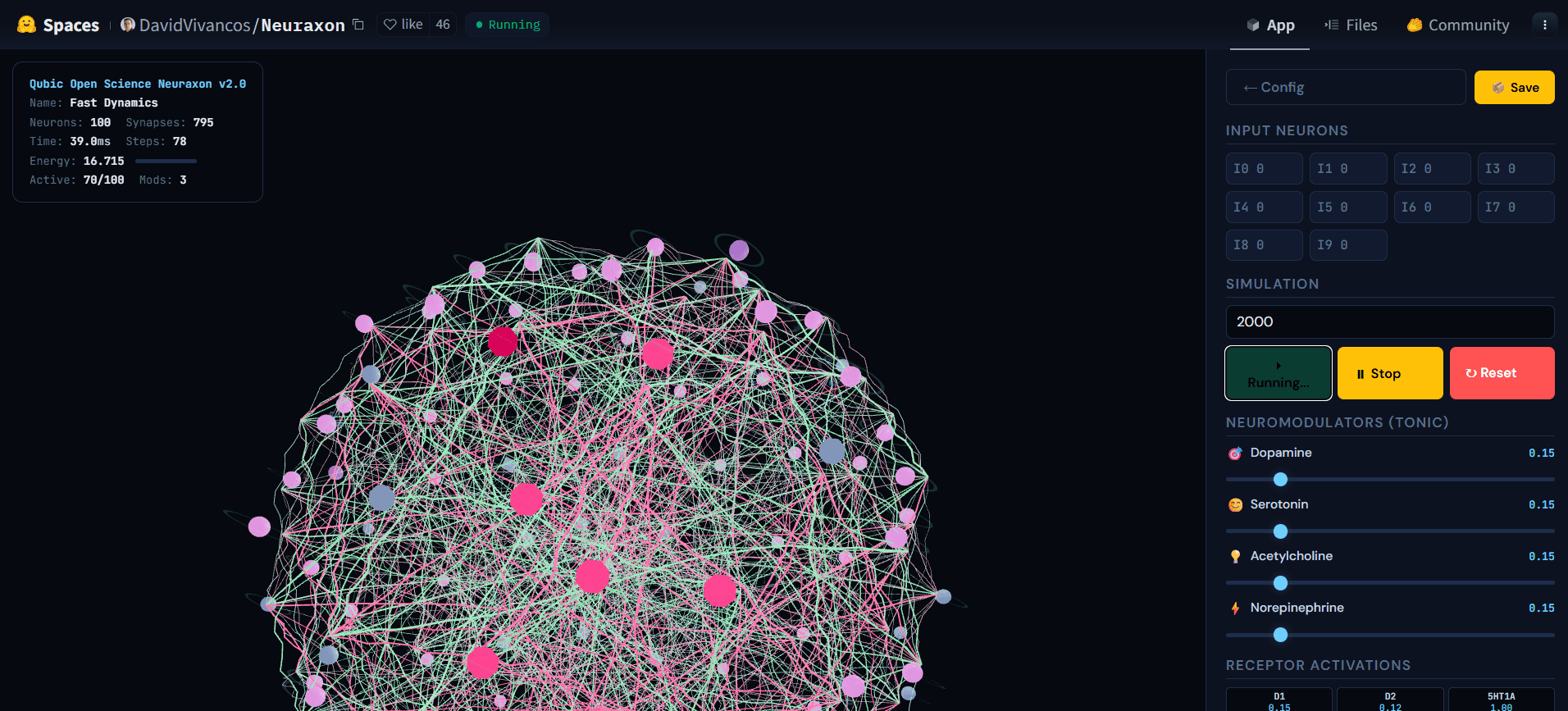
Gambar 5. Jaringan 3D Neuraxon selama simulasi aktif, menunjukkan aktivitas cascades di antara neuron keadaan ternary. Node yang sangat aktif (merah muda) menyebarkan sinyal melalui koneksi eksitatori (hijau) dan inhibitori (merah muda) sementara neuron lain tetap diam (abu-abu), menggambarkan rezim reverberasi dekat rasio cabang kritis (σ ≈ 1). Status seimbang ini — tidak sunyi maupun meledak — adalah apa yang Neuraxon terorganisir sendiri menggunakan prinsip kritisitas yang terinspirasi biologi. Jelajahi demo interaktif di huggingface.co/spaces/DavidVivancos/Neuraxon. Sumber: Tim Ilmiah Qubic.
Sebagai Jembatan Antara Observasi Neuroscientific dan Desain AI
Rasio cabang adalah salah satu dari sedikit magnitudo yang diukur dengan formalism yang sama dalam elektrofisiologi, fMRI, dan jaringan buatan. Ini memungkinkan pengujian hipotesis bidirectional: jika intervensi meningkatkan kritisitas biologis, kita dapat bertanya apakah intervensi yang sama — diterjemahkan ke dalam arsitektur buatan — meningkatkan komputasi model, dan sebaliknya. Prinsip ini menjadi sentral dalam kerangka neuromodulasi dan mekanisme penguncian astrositik yang telah kami kembangkan di volume akademi sebelumnya.
Sebagai Kriteria Fungsional, Bukan Estetika, untuk AI Terinspirasi Otak
Kritisitas adalah batasan operasional dengan konsekuensi empiris. Beroperasi dekat dengan rezim reverberasi meningkatkan — seperti yang diukur dalam evaluasi internal kami dan publikasi yang diajukan — kapasitas generalisasi, stabilitas di bawah gangguan input, kekayaan representasional, dan koherensi temporal dari penalaran. Efek ini secara kualitatif cocok dengan yang dilaporkan dalam literatur biologis (Cocchi et al., 2017) dan buatan (Cramer et al., 2020; Morales et al., 2023).
Rasio Cabang: Dari Fisika Statistik ke Arsitektur AI Terinspirasi Otak
Rasio cabang adalah salah satu dari raritas konseptual: cukup sederhana untuk diringkas menjadi satu formula, cukup dalam untuk menjembatani fisika statistik, ilmu saraf, AI, dan desain sistem. Untuk otak biologis, σ ≈ 1 tampaknya menjadi rezim di mana kombinasi virtuous dari sensitivitas, memori, ekspresivitas, dan ketahanan muncul. Untuk jaringan buatan, perbatasan yang sama — dijenamakan sebagai tepi kekacauan — memprediksi kapasitas komputasi maksimal.
Dan untuk Neuraxon, ini adalah prinsip panduan desain yang terinspirasi biologi: metrik yang dapat diaudit, mengatur diri sendiri, dan bermakna secara biologis yang membantu kami menjaga sistem tetap hidup, dalam pengertian terluas dari kata tersebut.
Referensi
Beggs, J. M., & Plenz, D. (2003). Longsor neuronal dalam sirkuit neokortikal. The Journal of Neuroscience, 23(35), 11167–11177. https://doi.org/10.1523/JNEUROSCI.23-35-11167.2003
Bertschinger, N., & Natschläger, T. (2004). Komputasi waktu nyata di tepi kekacauan dalam jaringan neural berulang. Neural Computation, 16(7), 1413–1436. https://doi.org/10.1162/089976604323057443
Boedecker, J., Obst, O., Lizier, J. T., Mayer, N. M., & Asada, M. (2012). Pemrosesan informasi dalam jaringan echo state di tepi kekacauan. Teori dalam Biosains, 131(3), 205–213. https://doi.org/10.1007/s12064-011-0146-8
Bornholdt, S., & Röhl, T. (2003). Jaringan neural kritis yang terorganisir sendiri. Physical Review E, 67(6), 066118. https://doi.org/10.1103/PhysRevE.67.066118
Cocchi, L., Gollo, L. L., Zalesky, A., & Breakspear, M. (2017). Kritisitas di otak: Sebuah sintesis neurobiologi, model dan kognisi. Kemajuan dalam Neurobiologi, 158, 132–152. https://doi.org/10.1016/j.pneurobio.2017.07.002
Cramer, B., Stöckel, D., Kreft, M., Wibral, M., Schemmel, J., Meier, K., & Priesemann, V. (2020). Kontrol kritisitas dan komputasi dalam jaringan neuromorfik spiking dengan plastisitas. Nature Communications, 11, 2853. https://doi.org/10.1038/s41467-020-16548-3
de Carvalho, J. X., & Prado, C. P. C. (2000). Kritisitas terorganisir sendiri dalam model Olami-Feder-Christensen. Physical Review Letters, 84(17), 4006–4009. https://doi.org/10.1103/PhysRevLett.84.4006
Derrida, B., & Pomeau, Y. (1986). Jaringan acak automata: Sebuah pendekatan sederhana yang dikeraskan. Europhysics Letters, 1(2), 45–49. https://doi.org/10.1209/0295-5075/1/2/001
Hagemann, A., Wilting, J., Samimizad, B., Mormann, F., & Priesemann, V. (2021). Menilai kritisitas dalam aktivitas neuron tunggal pra-kejang dari korteks epilepsi manusia. PLOS Computational Biology, 17(3), e1008773. https://doi.org/10.1371/journal.pcbi.1008773
Haldeman, C., & Beggs, J. M. (2005). Cabang kritis menangkap aktivitas dalam jaringan neural hidup dan memaksimalkan jumlah keadaan metastabil. Physical Review Letters, 94(5), 058101. https://doi.org/10.1103/PhysRevLett.94.058101
Hsu, D., Chen, W., Hsu, M., & Beggs, J. M. (2008). Sebuah hipotesis terbuka: Apakah epilepsi dipelajari, dan bisakah itu dilupakan? Epilepsi & Perilaku, 13(3), 511–522. https://doi.org/10.1016/j.yebeh.2008.05.007
Langton, C. G. (1990). Komputasi di tepi kekacauan: Transisi fase dan komputasi yang muncul. Physica D: Nonlinear Phenomena, 42(1–3), 12–37. https://doi.org/10.1016/0167-2789(90)90064-V
Levina, A., Herrmann, J. M., & Geisel, T. (2007). Sinaps dinamis yang menyebabkan kritisitas terorganisir sendiri dalam jaringan neural. Nature Physics, 3(12), 857–860. https://doi.org/10.1038/nphys758
Magnasco, M. O. (2022). Ketahanan dan fleksibilitas fungsi neural melalui kritisitas dinamis. Entropi, 24(5), 591. https://doi.org/10.3390/e24050591
Meisel, C., Klaus, A., Vyazovskiy, V. V., & Plenz, D. (2017). Interaksi antara korelasi temporal jarak jauh dan dekat membentuk dinamika korteks di berbagai keadaan kewaspadaan. The Journal of Neuroscience, 37(42), 10114–10124. https://doi.org/10.1523/JNEUROSCI.0448-17.2017
Morales, G. B., di Santo, S., & Muñoz, M. A. (2023). Mengungkap dinamika intrinsik jaringan neural biologi dan buatan: Dari kritisitas ke representasi optimal. Frontiers in Complex Systems, 1, 1276338. https://doi.org/10.3389/fcpxs.2023.1276338
Plenz, D., Ribeiro, T. L., Miller, S. R., Kells, P. A., Vakili, A., & Capek, E. L. (2021). Kritisitas yang terorganisir sendiri di otak. Frontiers in Physics, 9, 639389. https://doi.org/10.3389/fphy.2021.639389
Shew, W. L., Yang, H., Petermann, T., Roy, R., & Plenz, D. (2009). Longsor neuronal menyiratkan rentang dinamis maksimum dalam jaringan kortikal pada kritisitas. The Journal of Neuroscience, 29(49), 15595–15600. https://doi.org/10.1523/JNEUROSCI.3864-09.2009
Shew, W. L., Yang, H., Yu, S., Roy, R., & Plenz, D. (2011). Kapasitas informasi dan transmisi dimaksimalkan dalam jaringan kortikal seimbang dengan longsor neuronal. The Journal of Neuroscience, 31(1), 55–63. https://doi.org/10.1523/JNEUROSCI.4637-10.2011
Spitzner, F. P., Dehning, J., Wilting, J., Hagemann, A., Neto, J. P., Zierenberg, J., & Priesemann, V. (2021). MR. Estimator, sebuah alat untuk menentukan skala waktu intrinsik dari aktivitas spiking yang disampling. PLOS ONE, 16(4), e0249447. https://doi.org/10.1371/journal.pone.0249447
Timme, N. M., Marshall, N. J., Bennett, N., Ripp, M., Lautzenhiser, E., & Beggs, J. M. (2016). Kritisitas memaksimalkan kompleksitas dalam jaringan neural. Frontiers in Physiology, 7, 425. https://doi.org/10.3389/fphys.2016.00425
Turrigiano, G. G. (2008). Neuron penyesuaian diri: Skala sinaptik dari sinaps excitatory. Cell, 135(3), 422–435. https://doi.org/10.1016/j.cell.2008.10.008
Wang, J., Cao, R., Brunton, B. W., Smith, R. E. W., Buckner, R. L., & Liu, T. T. (2025). Kontribusi genetik terhadap kritisitas otak dan hubungannya dengan fungsi kognitif manusia. Proceedings of the National Academy of Sciences, 122(26), e2417010122. https://doi.org/10.1073/pnas.2417010122
Wilting, J., Dehning, J., Pinheiro Neto, J., Rudelt, L., Wibral, M., Zierenberg, J., & Priesemann, V. (2018). Beroperasi dalam rezim reverberasi memungkinkan penyetelan cepat kondisi jaringan sesuai kebutuhan tugas. Frontiers in Systems Neuroscience, 12, 55. https://doi.org/10.3389/fnsys.2018.00055
Wilting, J., & Priesemann, V. (2018). Menyimpulkan keadaan dinamis kolektif dari sistem yang jarang diamati. Nature Communications, 9, 2325. https://doi.org/10.1038/s41467-018-04725-4
Wilting, J., & Priesemann, V. (2019). 25 tahun kritisitas dalam ilmu saraf — Hasil yang telah ditetapkan, kontroversi terbuka, konsep baru. Current Opinion in Neurobiology, 58, 105–111. https://doi.org/10.1016/j.conb.2019.08.002
Yu, C. (2022). Menuju analisis terpadu dari hipotesis kritisitas otak: Meninjau beberapa alat yang tersedia. Frontiers in Neural Circuits, 16, 911245. https://doi.org/10.3389/fncir.2022.911245
Zeraati, R., Engel, T. A., & Levina, A. (2024). Memperkirakan skala waktu intrinsik dan kritisitas dari rekaman neural: Metode dan jebakan. Current Opinion in Neurobiology, 86, 102871. https://doi.org/10.1016/j.conb.2024.102871
Zimmern, V. (2020). Mengapa kritisitas otak relevan secara klinis: Tinjauan skoping. Frontiers in Neural Circuits, 14, 54. https://doi.org/10.3389/fncir.2020.00054
Jelajahi Akademi Kecerdasan Neuraxon secara Lengkap
Ini adalah Volume 8 dari Akademi Kecerdasan #Neuraxon oleh Tim Ilmiah #Qubic . Jika Anda baru bergabung dengan kami, jelajahi seluruh seri untuk membangun pemahaman penuh tentang ilmu di balik Neuraxon, #aigarth , dan pendekatan Qubic terhadap kecerdasan buatan #decentralized yang terinspirasi otak:
NIA Vol. 1: Mengapa Kecerdasan Tidak Dihitung Dalam Langkah, tetapi Dalam Waktu — Menjelajahi mengapa kecerdasan biologis beroperasi dalam waktu kontinu daripada langkah komputasi diskrit seperti LLM tradisional.
NIA Vol. 2: Dinamika Ternary sebagai Model Kecerdasan Hidup — Menjelaskan dinamika ternary dan mengapa logika tiga status (excitatory, netral, inhibitory) penting untuk memodelkan sistem hidup.
NIA Vol. 3: Neuromodulasi dan AI Terinspirasi Otak — Membahas neuromodulasi dan bagaimana sinyal kimia otak (dopamin, serotonin, asetilkolin, norepinefrin) menginspirasi arsitektur Neuraxon.
NIA Vol. 4: Jaringan Neural dalam AI dan Ilmu Saraf — Perbandingan mendalam antara jaringan neural biologis, jaringan neural buatan, dan pendekatan jalur ketiga Neuraxon.
NIA Vol. 5: Astrocytes dan AI Terinspirasi Otak — Bagaimana penguncian astrositik mengubah plastisitas jaringan neural melalui kerangka AGMP di Neuraxon.
NIA Vol. 6: Mesin Sadar vs Organisme Cerdas: Kesadaran AI Dijelaskan — Menjelajahi kesadaran AI melalui lensa Teori Tempat Kerja Global, Teori Informasi Terintegrasi, dan pengkodean prediktif.
NIA Vol. 7: Permainan Kehidupan Conway, Kehidupan Buatan, dan Ekosistem Digital — Ilmu di balik Qubic, Aigarth, dan pendekatan Neuraxon terhadap kompleksitas yang muncul dan kritisitas terorganisir sendiri dalam AI terdesentralisasi.
Qubic adalah jaringan terdesentralisasi, sumber terbuka untuk teknologi eksperimental. Untuk mempelajari lebih lanjut, kunjungi qubic.org. Bergabunglah dalam diskusi di X, Discord, dan Telegram.