Написано командой Qubic Scientific
 Академия интеллекта Neuraxon — Том 3
Академия интеллекта Neuraxon — Том 3

1. Нейромодуляция в мозге: основа адаптивного интеллекта
Нейромодуляция относится к набору механизмов, которые регулируют, как нервная система функционирует в любой данный момент, не изменяя ее базовую архитектуру. Благодаря нейромодуляции мозг может учиться быстро или медленно, становиться исследовательским или консервативным и оставаться открытым к новизне или сосредоточиваться на том, что уже известно. Провода не меняются; меняется только способ, которым эти провода используются. Эта концепция является центральной для понимания искусственного интеллекта, вдохновленного мозгом, и архитектуры, стоящей за Neuraxon от Qubic.
Ионотропные против метаботропных рецепторов: две временные шкалы нейронной сигнализации
Чтобы правильно понять нейромодуляцию, важно различать две формы химического действия в мозге. С одной стороны, существуют нейромедиаторы, которые действуют на ионотропные рецепторы, такие как глутамат и ГАМК. Эти рецепторы являются ионными каналами: когда они активируются, они вызывают немедленные электрические изменения в нейроне, на уровне миллисекунд. Это соответствует быстрому уровню нейронных вычислений: передается конкретная информация, интегрируются сенсорные сигналы, принимаются быстрые решения, и генерируется нейронная активность, которая поддерживает восприятие, движение и мышление в реальном времени.
С другой стороны, существуют нейромедиаторы, такие как дофамин, норэпинефрин, серотонин и ацетилхолин, чье основное действие осуществляется через метаботропные рецепторы. Эти рецепторы не генерируют электрический сигнал напрямую. Вместо этого они активируют внутриклеточные сигнальные каскады, которые изменяют внутренние свойства нейрона на более длительные периоды времени, секунды, минуты или более. Это представляет собой медленный динамический уровень нейронной обработки, который является основополагающим для того, как мозг адаптируется и учится.
Интуитивный способ подумать об этом различии — это метафора морского порта. Ионотропные рецепторы похожи на пловцов, серферов или маленькие лодки, которые быстро входят и выходят. Метаботропные рецепторы, напротив, аналогичны большим грузовым кораблям. Для их швартовки нужны разрешения, требуется координация, и логистика порта должна быть отрегулирована. Эти метаботропные рецепторы изменяют синаптическую пластичность и легкость, с которой нейрон реагирует — эта медленная модуляция не передает информацию, а скорее изменяет внутренние правила системы.
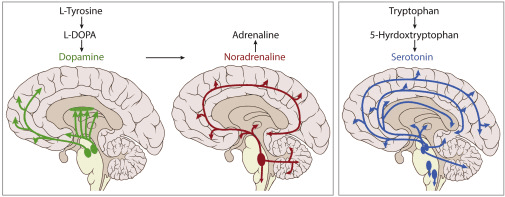
Четыре нейромодулятора: дофамин, норэпинефрин, серотонин и ацетилхолин
Здесь вступают в игру основные нейромодуляторные системы. Каждый из этих четырех нейротрансмиттеров играет свою уникальную роль в регулировании того, как мозг обрабатывает информацию, учится и адаптируется:
Дофамин, в основном исходящий из вентральной тегментальной области и черной субстанции, не сигнализирует о удовольствии как таковом, а скорее, когда что-то имеет значение для обучения. Он регулирует чувствительность системы к ошибкам и новизне. Как показал Шульц (2016) в своей основополагающей работе о кодировании ошибки предсказания вознаграждения дофамином, дофамин сигнализирует о разнице между ожидаемыми и фактическими результатами, что является критически важным механизмом для обучения с подкреплением как в биологических, так и в искусственных системах.
Норэпинефрин (норэпинефрин), выделяющийся в основном из латерального покрышечного ядра, регулирует возбуждение и баланс между исследованием и эксплуатацией. Когда его тон высок, мозг становится более чувствительным к неожиданным изменениям и менее привязанным к рутинным действиям. Это согласуется с интегративной теорией, предложенной Астон-Джонсом и Коэном (2005), которая связывает функцию латерального покрышечного ядра — норэпинефрина с адаптивным контролем прибыли и принятием решений в условиях неопределенности.
Серотонин, исходящий из ядер рафе, модулирует настроение, сон, торможение и стабильность поведения. Как исследовано в работе Даяна и Хуиса (2009), серотонин не заставляет систему учиться быстро, а скорее ждать, избегать импульсивных реакций и поддерживать поведение, когда окружающая среда неустойчива. Он играет критическую роль в терпении и долгосрочном планировании.
Ацетилхолин, выделяющийся из ядер базального переднего мозга в стволе мозга, играет центральную роль в внимании и обучении, зависящем от контекста. Он облегчает открытие корковых сетей для релевантной сенсорной информации и обеспечивает синаптическую пластичность, когда окружающая среда этого требует. Он особенно важен, когда необходимо выучить что-то новое, и поэтому является необходимым для адаптивных нейронных вычислений.
Благодаря этому комбинированному действию один и тот же стимул может вызывать разные реакции в зависимости от нейромодуляторного состояния. Цепь остается той же, но способ ее работы изменился. Именно поэтому мозг не реагирует одинаково, когда он внимателен, как когда он устал, и не обучается одинаково в рутинных ситуациях, как в условиях новизны или неожиданности.
Мета уровень: окна пластичности и адаптивное обучение
Существует также третий, более глубокий уровень, который можно понять как мета уровень нейронной регуляции. Этот уровень не регулирует непосредственно нейронную активность или ее скорость, а скорее условия, при которых система может измениться устойчиво. В мозге совпадение активности между нейронами не гарантирует обучение. Чтобы связь укрепилась или ослабла, нейромодуляторное состояние должно это позволить. Это как если бы был тихий сигнал, говорящий: "сейчас да" или "сейчас нет".
Таким образом, нейромодуляция действует как система, которая открывает или закрывает окна пластичности, решая, когда ошибка, опыт или совпадение заслуживает консолидации. Эта многомасштабная архитектура, быстрая, медленная и мета, существует потому, что интеллектуальная система не может всегда применять одни и те же правила. Как объяснила Мардэр (2012) в своем знаковом обзоре, нейромодуляция нейронных цепей — это то, как мозг достигает поведенческой гибкости, не перестраивая свою архитектуру.
Состояние тела, уровень энергии, усталость или боль являются частью внутренней среды. Новизна, угроза, возможность, повторение или предсказуемость являются частью внешней среды. Нейромодуляторные системы переводят эти условия в функциональные состояния. Через дофамин, норадреналин, серотонин и ацетилхолин мозг оценивает, заслуживает ли ситуация обучения, требуется ли осторожность, предпочтительнее ли исследование или сохранение, и является ли ошибка информативной или просто шумом. Среда не диктует непосредственно реакцию, но модулирует правила, по которым мозг реагирует. Этот принцип лежит в основе того, что Фристон (2010) описал как принцип свободной энергии, единая структура, предполагающая, что мозг постоянно минимизирует удивление через адаптивные внутренние модели.
2. Почему большие языковые модели и архитектуры трансформеров не имеют нейромодуляции
Большие языковые модели (LLM) и архитектуры на основе трансформеров не обладают нейромодуляцией. Хотя они обрабатывают длинные последовательности и достигли выдающихся результатов в обработке естественного языка, им не хватает системы, которая динамически регулирует режим работы модели во время вывода.
Статическая природа систем ИИ на основе трансформеров
Обучение в LLM происходит в фазах обучения, которые полностью отделены от использования. Веса корректируются через обратное распространение ошибки, и после завершения обучения модель переходит в фиксированное состояние. Во время вывода нет пластичности и нет стойких изменений в зависимости от контекста. Система не решает, когда уместно учиться, а когда следует стабилизироваться, потому что она не учится во время работы. Это основное ограничение, которое подтвердили недавние исследования, LLM не имеют истинных внутренних моделей мира и способности адаптироваться в реальном времени.
Некоторые подходы, вдохновленные нейромодуляцией, пытаются приблизить определенные эффекты, регулируя такие параметры, как скорость обучения во время тренировки, активируя или деактивируя подсети или модулируя функции активации. Однако это всего лишь внешние оптимизации, а не внутренние системы, которые регулируют активность и пластичность в реальном времени. Как утверждали Мэй, Мюллер и Рамасвами (2022) в журнале Trends in Neurosciences, информирование глубоких нейронных сетей многоуровневыми принципами нейромодуляторных систем остается открытой задачей, которую текущие архитектуры LLM не решили.
Хотя нейромодуляция иногда упоминается в контексте ИИ, LLM и трансформеры остаются частичными приближениями, а не системами, сопоставимыми с мозгом. Разрыв между статическими матричными вычислениями и динамическим, зависимым от состояния регулированием, обнаруживаемым в биологических нейронных сетях, именно то, что делает архитектуры ИИ, вдохновленные мозгом, такими как Neuraxon, необходимым следующим шагом к адаптивному искусственному интеллекту.
3. Как Neuraxon вычисляет нейромодуляцию: архитектура ИИ, вдохновленная мозгом
В Neuraxon вычисление — это процесс, который разворачивается в непрерывное время. Код выражает систему, которая поддерживает внутренние состояния, s(t), которые развиваются даже при отсутствии четких внешних стимулов. Эти состояния влияют на будущее поведение, создавая живую нейронную систему, которая всегда активна, концепция, подробно исследованная в исследовательской работе Neuraxon.
Быстрая, медленная и мета динамика в нейронных вычислениях
Neuraxon явно включает в себя быстрые, медленные и мета динамики, отражая многомасштабную временную архитектуру, найденную в биологическом мозге. Быстрая динамика управляет немедленной передачей активности, аналогично быстрому нейронному сигналу через ионотропные рецепторы. Медленная динамика вводит накопление, постоянство и стабилизацию паттернов, позволяя системе сохранять информацию за пределами мгновения, подобно тому, как метаботропные рецепторы модулируют нейронную функцию на протяжении секунд и минут. Мета динамика воздействует на правила взаимодействия между первыми, модулируя, когда система становится более чувствительной к изменениям и когда она стремится сохранить свое состояние.
Нейромодуляция в Neuraxon не реализуется как внешняя настройка параметров. Система не принимает решения о том, что учить, а скорее о том, при каких условиях она может изменяться. Это отражает то, как биологические нейромодуляторы, такие как дофамин и серотонин, создают окна пластичности, а не напрямую кодируют информацию. Вы можете исследовать эти динамики на практике с помощью интерактивной 3D-симуляции Neuraxon на HuggingFace Spaces, где вы можете регулировать уровни дофамина, серотонина, ацетилхолина и норэпинефрина в реальном времени и наблюдать, как они влияют на поведение сети.
От биологических принципов к децентрализованному ИИ
Этот подход не воспроизводит молекулярную или анатомическую сложность мозга, что в настоящее время невозможно воспроизвести. Нет тысяч рецепторов или реальных биологических сетей. Однако он сохраняет и вычисляет важный принцип: интеллект является адаптивным и, следовательно, требует внутренних динамик, состояния и модуляции.
Архитектура нейромодуляции Neuraxon является ключевой частью более широкой концепции Qubic для децентрализованного ИИ. Интегрируя Neuraxon с эволюционной структурой Aigarth Intelligent Tissue, Qubic создает систему, где миллионы архитектур на основе Neuraxon могут развиваться, конкурировать и улучшаться через распределенные вычисления, поддерживаемые механизмом консенсуса Qubic network’s Useful Proof of Work (UPoW).
4. Исследуйте нейромодуляторы с помощью интерактивной демонстрации Neuraxon
Хотите узнать, как нейромодуляция работает в системе ИИ, вдохновленной мозгом? Демонстрация Neuraxon Mood Mixer позволяет вам регулировать уровни дофамина, серотонина, ацетилхолина и норэпинефрина в реальном времени и наблюдать, как эти нейромодуляторы влияют на поведение нейронной сети. Это практический способ понять принципы, обсуждаемые в этой статье, и увидеть различие между статическими вычислениями ИИ и динамической, зависимой от состояния обработкой.
5. Математика, стоящая за многомасштабной нейромодуляцией Neuraxon
Темпоральные динамики в Neuraxon регулируются тремя дифференциальными уравнениями, которые захватывают быстрые, медленные и мета временные шкалы нейронных вычислений:

Здесь, τ_fast < τ_slow < τ_meta отражают их различные временные шкалы, при этом τ_meta значительно больше, чтобы захватить «ультрамедленный» характер метаботропных эффектов. Эта математическая структура напрямую реализует биологический принцип, согласно которому нейромодуляция работает на гораздо более медленных временных шкалах, чем быстрая синаптическая передача, как описано Нортхоффом и Хуаном (2017) в их работе о том, как временные динамики мозга медируют сознание.
Научные ссылки
Даян, П., и Хуис, Q. J. M. (2009). Серотонин, торможение и негативное настроение. PLoS Computational Biology.
Мардэр, Э. (2012). Нейромодуляция нейронных цепей: назад в будущее. Нейрон.
Шульц, В. (2016). Кодирование ошибки предсказания вознаграждения дофамином. Диалоги в клинической нейробиологии.
Астон-Джонс, Г., и Коэн, J. D. (2005). Интегративная теория функции латерального покрышечного ядра — норэпинефрина. Ежегодный обзор нейробиологии.
Мэй, Л., Мюллер, Э., и Рамасвами, С. (2022). Информирование глубоких нейронных сетей многоуровневыми принципами нейромодуляторных систем. Trends in Neurosciences.
Фристон, К. (2010). Принцип свободной энергии: единая теория мозга? Nature Reviews Neuroscience.
Нортхофф, Г., и Хуан, З. (2017). Как временные и пространственные аспекты мозга медируют сознание и его расстройства? Сознание и когнитивные процессы, 57, 1–10.